CORSO DI ALFABETIZZAZIONE IN IMMUNOLOGIA Parte 2 del Dott. Vincenzo D. Esposito
Il linguaggio molecolare dell’immunità: citochine, recettori, chemochine e sistema maggiore di istocompatibilità spiegati in modo chiaro e rigoroso
CAPITOLO 4 – Citochine
Codici di comunicazione all’interno del sistema dell’immunità innata
Grazie a queste molecole, le cellule adattano il loro comportamento ai cambiamenti che si verificano nel microambiente in risposta all’ingresso di un elemento estraneo.
Esistono recettori capaci di catturare questi segnali: alcuni sono costitutivamente presenti e attivi sulle membrane cellulari, altri sono inducibili, cioè vengono espressi solo in risposta al segnale ricevuto o durante specifiche fasi del ciclo cellulare.
I principali codici di comunicazione sono:
- il sistema delle citochine e delle chemochine
- il sistema maggiore di istocompatibilità (MHC)
- gli ormoni
Le cellule dell’immunità innata secernono citochine nel microambiente, e l’informazione che esse portano viene percepita solo dalle cellule circostanti che possiedono il recettore corrispondente. La conseguenza è che tali cellule cambiano comportamento dopo l’interazione citochina–recettore.
Le citochine hanno quindi una selettività: nello stesso microambiente possono essere presenti cellule che non esprimono il recettore specifico e che quindi non ricevono il messaggio.
Il linguaggio delle cellule dell’immunità innata è composto da due grandi famiglie:
- Interleuchine propriamente dette, che creano connessioni tra cellule
- Chemochine, che attivano il movimento dei leucociti (dal greco kineo, muovere)
Le interleuchine sono indicate con numeri progressivi in base al momento della loro scoperta o alla prima attività biologica identificata.
Caratteristiche molecolari di interleuchine e chemochine
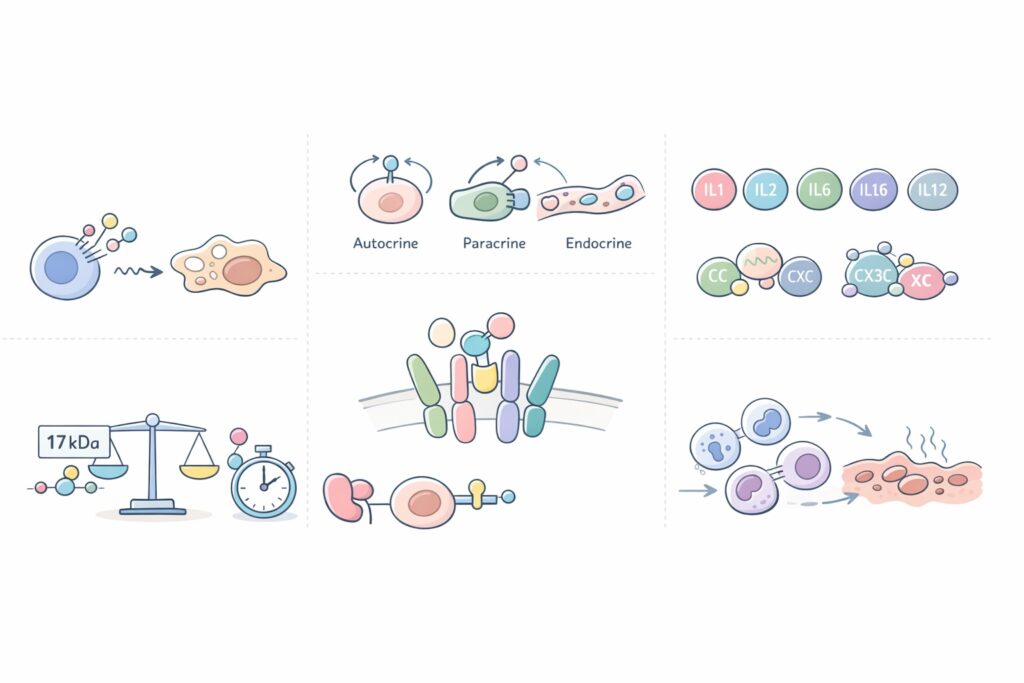
- peso molecolare di circa 17 kDa
- struttura in monomeri, dimeri o trimeri di proteine globulari
- emivita di pochi minuti
- neutralizzabili da inibitori specifici presenti nel siero
- rapidamente catabolizzate dal rene
Queste caratteristiche spiegano perché, nella maggior parte dei casi, le citochine non diffondono sistemicamente come gli ormoni, ma raggiungono solo le cellule vicine: sono prodotte in piccole quantità, vengono inattivate rapidamente e sono degradate dai reni.
Tipi di secrezione
- Secrezione polarizzata: la molecola viene rilasciata nello spazio tra cellula secernente e cellula ricevente.
- Secrezione autocrina: la cellula utilizza la citochina che essa stessa ha prodotto.
- Secrezione endocrina: la molecola agisce a distanza in tutto il corpo.
- Esempio: IL‑6 e IL‑1, prodotte nell’area infetta, raggiungono l’ipotalamo e inducono febbre.
Recettori per le citochine
- la loro espressione cambia durante la maturazione delle cellule bianche
- alcuni recettori legano la citochina ma non trasducono il segnale
- altri sono decodificanti, cioè trasmettono l’informazione all’interno della cellula
I recettori sono proteine globulari costituite da due o tre catene:
- catena alfa (CD25)
- catena beta (CD131)
- catena gamma (CD123)
La catena gamma è la più importante: una mutazione nel gene che la codifica provoca una grave immunodeficienza. Il gene si trova sul cromosoma X.
Il recettore per l’interleuchina 2 (IL‑2)
È un recettore predisposto a interagire con interleuchine costituite da due o tre catene. La sua conformazione varia durante l’attivazione cellulare, determinando la selettività del messaggio.
- La catena alfa (CD25) è la prima a essere espressa: indica che il linfocita è stato pre‑attivato.
- Le catene beta e gamma sono le uniche capaci di trasdurre il segnale al DNA.
La catena gamma segnala IL‑2 ma non la lega. La catena beta lega IL‑2 ma con bassa affinità. Solo un’alta concentrazione di IL‑2 permette il legame stabile.
Una volta legata, la catena beta cambia conformazione e interagisce con la catena gamma, formando un recettore trimerico ad alta affinità (alfa‑beta‑gamma). Questo complesso attiva i fattori di trascrizione che raggiungono il nucleo e regolano:
- ciclo cellulare
- proliferazione
- formazione di cloni linfocitari
Chemochine
Esistono due grandi categorie:
- Chemochine chemiotattiche: guidano le cellule sentinella verso il sito dell’infezione.
- Chemochine infiammatorie: reclutano cellule bianche durante la risposta immunitaria.
Sono classificate in quattro famiglie: CC, CXC, CX3C, XC, caratterizzate dalla presenza dell’amminoacido cisteina, che favorisce i legami molecolari.
Le chemochine infiammatorie stimolano le cellule linfatiche a produrre integrine, che permettono alle cellule sentinella di uscire dai capillari e raggiungere i tessuti infetti.
La loro interazione con i recettori specifici guida lo spostamento delle cellule verso il sito dell’infezione.
CAPITOLO 5 – Sistema maggiore di istocompatibilità (MHC)
Struttura e funzione
Gli antigeni di istocompatibilità sono i marcatori dell’individualità di ogni vertebrato, uomo compreso. Nell’uomo, i geni che ne determinano la produzione sono localizzati in una piccola regione del cromosoma 6.
Questi geni sono indicati con la sigla HLA (Human Leukocyte Antigens).
Esistono tre classi di antigeni di istocompatibilità:
Classe I e Classe II
Sono molecole transmembrana costituite da:
- una porzione citoplasmatica
- una porzione che attraversa la membrana
- una porzione extracellulare
Classe I Espresse su tutte le cellule nucleate, tranne che in bassa quantità su:
- cellule del sistema nervoso
- cellule endocrine
- cellule muscolari
- spermatozoi
L’infiammazione aumenta la loro espressione tramite interferone alfa e gamma.
Classe II Espresse costitutivamente su:
- cellule dendritiche
- cellule di Langerhans
- cellule epiteliali timiche
- cellule B
Possono essere indotte su:
- cellule endoteliali
- cellule epiteliali
- macrofagi
- neutrofili
- eosinofili
- fibroblasti
Funzioni principali
- Determinare l’identità biologica individuale
- Presentare frammenti proteici (antigeni) alle cellule T, attivando l’immunità adattativa
Gli antigeni di classe III non sono transmembrana ma molecole secrete, coinvolte nella fase di transizione tra immunità innata e adattativa.